咨询热线
18862190290
18862190290

膜蛋白是与细胞区室或细胞器的细胞膜相关或附着的蛋白质。它们代表了最大和最重要的蛋白质类别之一,可以分为外周或整体。
在过去的几十年里,已知的人类蛋白质编码基因的数量总是略有变化,但近年来,绝对数量围绕着20.20个基因(Piovesan,Antonaros和Vitale)旋转。 这些蛋白质中约有三分之一是分泌蛋白或膜结合蛋白,虽然这是整个蛋白质组的重要组成部分,但其中只有少数在结构上是已知的。由于所有获批的疗法中约有一半靶向膜蛋白,解析这些治疗相关膜蛋白的结构非常有利于未来的药物设计。
膜蛋白质组和分泌组被认为是最大和最重要的蛋白质类别之一。膜蛋白被定义为与细胞膜或细胞内细胞器相关或附着的蛋白质。它们分为外周蛋白和整体蛋白。外周膜蛋白是 时间上 与脂质双层相关,但不能全部跨越膜。通过外周区域的穿孔或与整合膜蛋白偶联来实现对脂质双层的附着(参见 图3,B&C).这些完整的蛋白质嵌入,跨越整个脂质双层,并包含位于膜内的疏水性α-螺旋或β-桶结构。根据它们的细胞功能,它们可以进一步细分为受体或通道等组。
与亲水性细胞外和细胞内结构域一起,大多数膜蛋白表现出两亲性特征。两亲性特征还产生了一种特征,通过该特征通常可以识别完整的膜蛋白。这是由于它们的一级结构在其线性序列中含有19-5582个疏水氨基酸,需要跨越膜的疏水内部。带有指向桶外部的疏水残基的β桶也可以作为膜蛋白的良好指标。
在许多生理和病理过程中,试剂、转录因子、蛋白质或离子需要通过膜屏障。如果成功,它们会触发信号通路,发送生长和凝血因子或传递无法穿过脂质双层的蛋白质信号,如细胞因子。因此,膜蛋白位于许多细胞过程的一个非常重要的交叉点。
为此,膜蛋白接管了许多关键功能,例如蛋白质和离子通过特殊通道的运输或许多生物体内的信号转导。它们还负责细胞间连接或细胞间识别,使细胞之间的快速通信和外来细胞的有效识别,这对免疫系统至关重要。
目前,TCDB(转运体分类数据库)列出了69个超家族,其中有1600多个转运蛋白家族。由于数量庞大,我们只想在下面对这些传输器类进行一个小的概述:
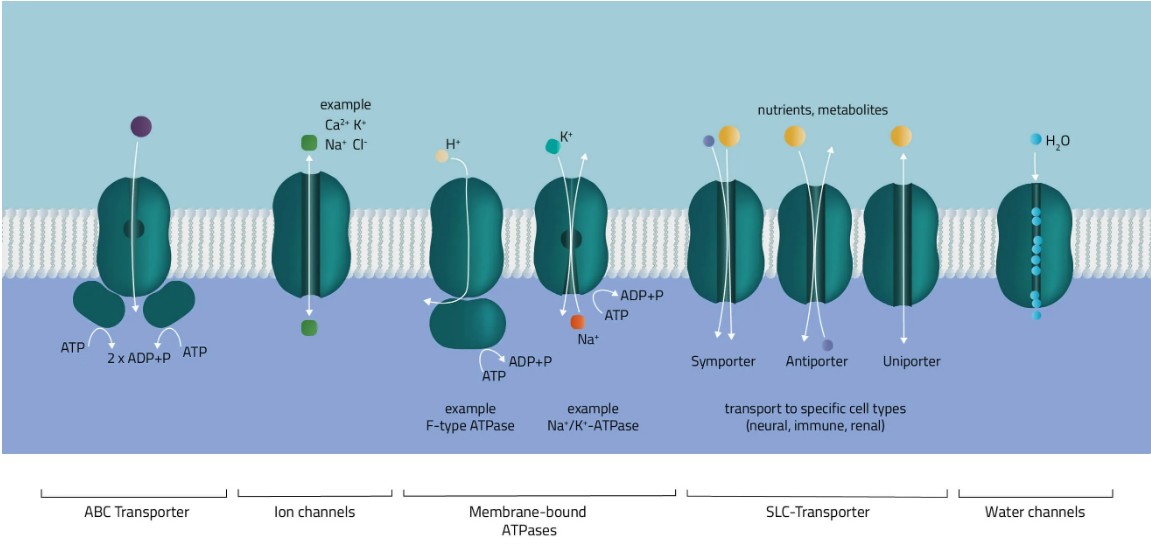
ABC-转运车
ATP结合盒转运蛋白超家族是最大的基因家族之一。在大多数情况下,它们由多个亚基组成,分为疏水跨膜结构域和亲水膜相关ATP酶。作为分子泵,它们利用ATP水解的能量在细胞膜上移动各种溶质(Jones和George,2002)。
离子通道
造孔蛋白促进离子流过细胞膜。根据类型进行细分,通过分类为门控机制,按离子类型或细胞定位进行细分。第一组中类型是电压门控离子通道,例如许多Na+, K+, 或 CA2+ 渠道。从生物学上讲,它们是神经系统的关键组成部分。同样,它们参与肌肉收缩或T细胞活化也同样重要。
膜结合ATP酶
顾名思义,ATP酶超家族利用ATP来执行其功能。它们分为 F、V 和 P 型 ATP 酶。F型和V型ATP酶被归类为旋转ATP酶,而P型ATP酶利用ATP水解释放的自由能驱动其构象变化(Palmgren和Nissen,2011;Pizzagalli, Bensimon, and Superti-Furga, 2020)。一种ATP酶是Na+/K+-交换剂,根据其浓度梯度泵送钠和钾,以维持细胞膜电位。
SLC-转运车
溶质载体蛋白是转运蛋白的超家族,通过膜转移多种溶质。这些包括糖、氨基酸、维生素或金属等分子(Hediger 等。, 2013).因此,它们是进入或离开细胞的主要调节剂之一,许多生理和细胞过程都依赖于它们(Pizzagalli、Bensimon 和 Superti-Furga,2020 年)。
水道
水通道蛋白或水通道促进水流过膜。因此,它们在维持水平衡方面发挥着基本作用。
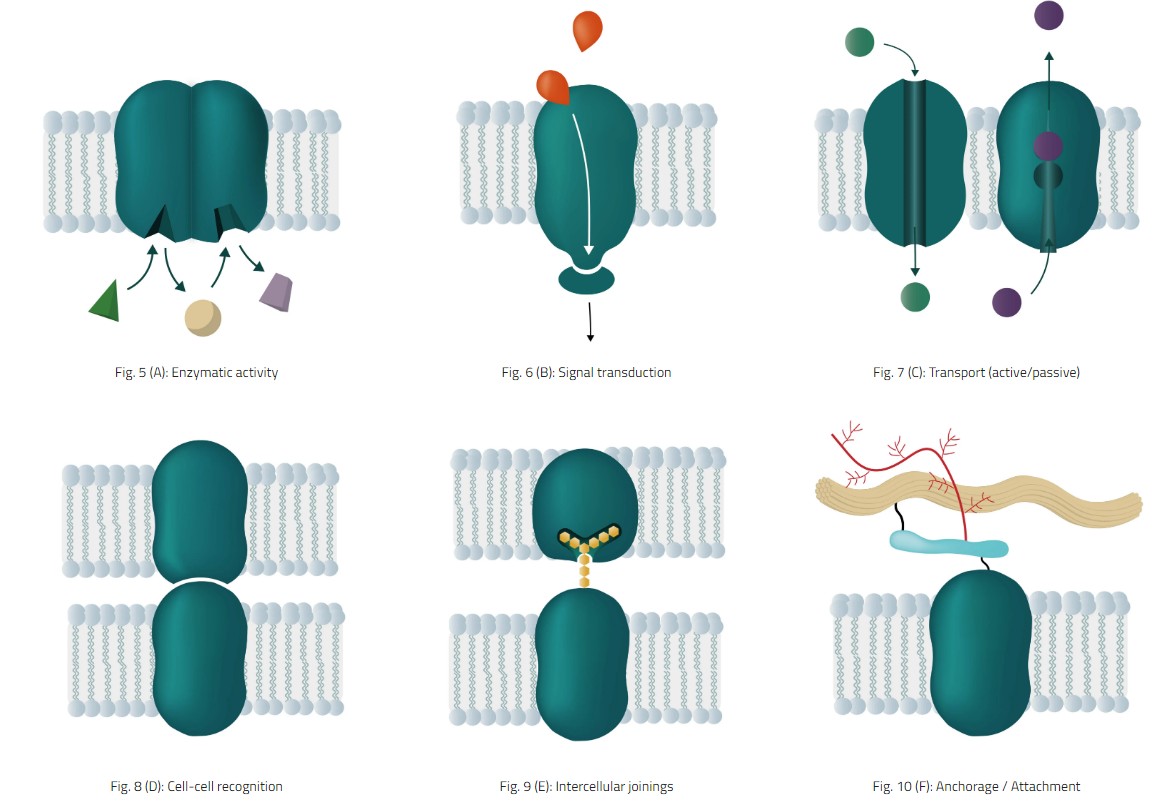
酶活性 - 各种代谢途径的代谢物和底物的加工
信号转导 - 化学信使与膜蛋白结合位点相互作用以发出信号
传输(主动/被动) - 在不同的细胞膜上移动分子和其他物质
细胞间识别 - 细胞之间的识别,即与免疫系统相关的细胞
细胞间连接 - 不同的结点,如间隙或紧密结连接相邻的电池
锚固/附件 - 对细胞骨架网络、蛋白质位置和某些形状的维持很重要
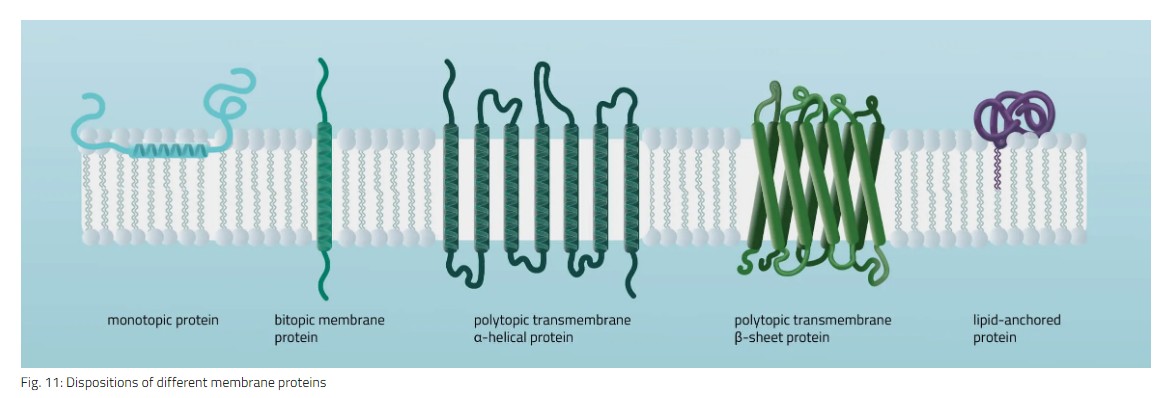
单主题 整体蛋白附着在膜双层的一侧。相互作用类型涉及例如平行于膜平面的两亲性α螺旋(参见 图11)或几个疏水环将蛋白质整体锚定。
双位 整体蛋白仅跨越脂质双层一次。典型的双位结构由跨膜结构域和两个细胞结构域(额外和内部)组成。
多主题 跨膜蛋白不止一次跨越脂质双层。跨膜结构域中α-螺旋和β-桶元素的不同组成是可能的(参见 图11 举两个例子)。
脂质锚定 蛋白质共价附着在磷脂双层中的脂质上。相互作用可以通过与膜脂质的共价键(脂化)或与膜脂质的静电/离子相互作用发生。
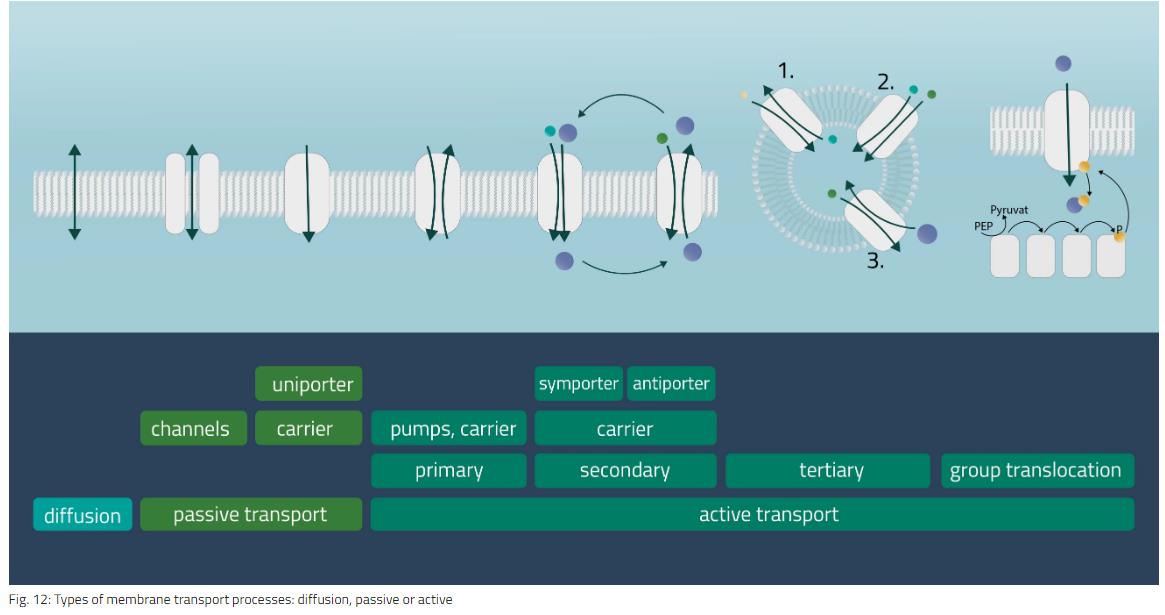
简单扩散
例如,分子、离子或颗粒从较高浓度的区域沿其梯度向下移动到较低浓度的区域。运动一直持续到达到平衡。带电粒子可以向一个方向或另一个方向移动这种膜电位。
被动运输
较大分子(如糖或氨基酸)的简单扩散的类似物。膜转运蛋白使在正常条件下无法通过膜的底物沿着浓度梯度向下移动。
通道蛋白
跨越细胞膜的特殊跨膜蛋白。门控机制可由配体、膜电位变化或机械过程(例如细胞骨架变化)触发。
载体蛋白
对于这种类型的被动转运,分子通过特殊载体的构象变化进行转移。这种变化是由基板对接到载体触发的。运输方式可以用单个分子(Uniport)进行,两个分子沿同一方向(Symport)或相反方向(Antiport)移动。
主动运输
主动运输是一种需要外部能量才能执行其机制的运输形式。这使得分子或离子能够相对于其浓度梯度或电势梯度传输。能量的形式可以是化学性质的(ATP)或电荷。也可以利用浓度梯度作为能量来源。
主要主动传输
质子和无机离子通过利用ATP的能量在细胞膜中移动。Na-K泵是这种传输形式的一个例子,它传递三个带正电荷的钠离子和两个同样带正电荷的钾离子。
二次主动传输
类似于被动载体蛋白同源蛋白和反转运蛋白,这次其中一个离子随着其浓度梯度移动,而第二个离子则与其相反移动。这可以在同一方向(Symport)或相反方向(Antiport)上实现。因此,电化学梯度是这里的驱动力。
三级主动运输
对于这种类型,三个转运体需要串联运行。第一转运蛋白建立分子的电化学梯度 一个 (主要活动)。第二转运蛋白利用分子 一个 为分子建立有利的电化学梯度 B (辅助活动)。最后,转运蛋白三利用分子梯度 B 移动分子 C 反对其浓度梯度(Hamm,Alpern和Preisig,2008)。
群体易位
这是一种特殊的细菌运输形式。要运输的基材在此过程中会发生化学变化。因此,不会产生浓度梯度。作为一种能量形式,可以使用ATP,但还有其他能量来源,如PEP(磷酸转移酶系统),也用于葡萄糖运输过程(见 图12、组易位)。
同时,已经应用的实验方法,如 纳米盘系统,环烷烃改性 两栖动物或 阿斯蒂 共聚物也将进一步发展,推动我们的知识边界向前发展。
未来,人工智能和此类实验方法的交织将引入生物和医学研究的新时代,塑造数字生物学或生物医学计算等领域。新方法将更新我们的药物发现过程,极大地加速它们。无论我们追求什么路径,膜蛋白质组都将在其中发挥至关重要的作用。
 扫一扫,关注微信
扫一扫,关注微信 当前位置:
当前位置: