咨询热线
18862190290
18862190290

“绿茶有助于对抗癌症!" - 一段时间以来,人们已经知道绿茶是如此健康,以至于它甚至改善了日本的癌症统计数据[1]。但这是为什么呢?这个问题的答案只能通过表观遗传学找到。术语“表观遗传学"由遗传学和表观发生学(即生物的发育)组成,描述了一个研究环境对我们基因的影响的研究领域[2]。所以问题是基因在什么情况下被打开,什么时候再次失活。基因活性的这些变化不是基于DNA序列的变化,例如通过突变或重组。相反,它们基于染色质或与DNA结合的蛋白质的化学变化。 虽然这些表观遗传印记无法在基因型中检测到,但由于DNA序列没有改变,它们仍然可以很好地在表型中观察到,也可以传递给子细胞[3]。
在绿茶的情况下,它是一种特定的表观遗传机制,触发抗癌作用:当非发酵茶叶被冲泡时,物质表没食子儿茶素-3-没食子酸酯(EGCG)被释放出来。然后EGCG重新激活编码抗癌蛋白的基因。该基因经常甲基化,特别是在老年人中,因此是沉默的。因此,当饮用绿茶时,该基因再次被打开,从而可以发挥其抗癌作用[2]。这只是表观遗传学的众多例子之一,这些例子显示了环境影响如何调节我们的基因。在许多科学家眼中,这个新兴的研究领域有望更好地了解疾病并对其治疗有新的了解。
基本的表观遗传机制之一是所谓的组蛋白的修饰。这些是基本蛋白质,作为染色质的组成部分,对DNA的包装至关重要。组蛋白H2A,H2B,H3和H4各两个拷贝形成八个组蛋白的蛋白质复合物。这种组蛋白八聚体形成核小体的核心,DNA包裹在其周围。这样的核小体代表了DNA的最小包装单位[4]。从它突出组蛋白链的末端,组蛋白链是组蛋白修饰酶的靶标(见图1)。翻译后修饰组蛋白的描述可以追溯到1960年代。当时,对小牛胸腺组蛋白的分析检测到甲基化赖氨酸,不久后,也可以检测到乙酰化赖氨酸[5,6]。今天 赖氨酸的甲基化和乙酰化代表了最著名的组蛋白修饰。然而,在组蛋白上发现了许多其他翻译后修饰(PTM),这些修饰在基因表达的调节中起重要作用(见图1)。
组蛋白修饰既可以发生在组蛋白的非结构化N端和C端,也可以发生在核小体核心内的球状区域(图1)。一个特定的命名法已经演变为描述不同的修饰:首先,给出组蛋白的名称(例如H3)。接下来是参与其单字母代码的氨基酸(例如K表示赖氨酸)以及氨基酸在蛋白质中的位置。现在提到了修饰的类型(例如,Me代表甲基,P代表磷酸盐或Ac代表乙酰基)。最后,还可以设定甲基的数量(在赖氨酸和精氨酸的情况下)[7]。例如,名称H3K4me3代表组蛋白3位置4处赖氨酸的三甲基化。
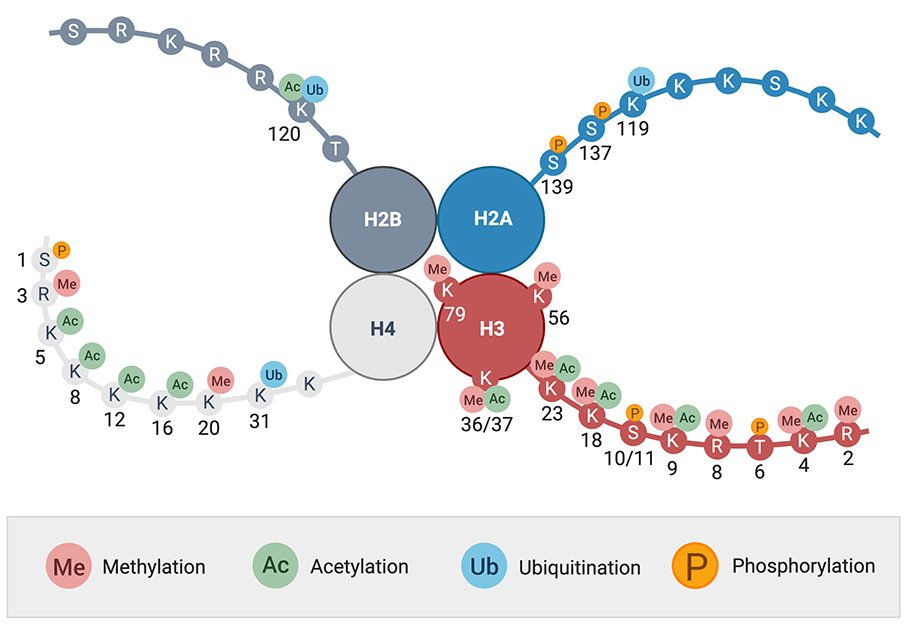
图1:组蛋白H2A、H2B、H3和H4的翻译后修饰.图中显示了四种主要的PTM甲基化,乙酰化,泛素化和磷酸化。它们发生在组蛋白的N端和C端以及核小体核心内的球状区域[8]。
在我们仔细研究个体翻译后组蛋白修饰及其对基因表达的影响之前,我们将更详细地讨论表观遗传学研究的基本方法之一:染色质免疫沉淀(ChIP)。该方法可用于分析完整细胞中蛋白质与特定DNA片段的相互作用[9]。目的是找出所检查的蛋白质是否与特定的基因区域相关[10]。ChIP可用于研究转录因子与启动子的结合,也可用于探索各种组蛋白修饰的分布,从而作为表观遗传基因调控的基础[11]。
基本原理是通过用甲醛固定来捕获存在于某个时间点的蛋白质-DNA结合(图2)。随后,从细胞中提取的DNA通过超声处理被片段化成50至3072个碱基对的片段,留下结合的蛋白质附着在DNA上。在下一步中,使用DNA相关蛋白特异性抗体选择性提取与目标蛋白相关的DNA片段。随后,分离的DNA-蛋白质复合物通过热处理再次溶解(图2)。游离的DNA片段现在可以纯化,并且使用进一步的方法(例如PCR,NGS)可以定量和鉴定。由此可以得出结论,该蛋白质是否与活细胞中的相关DNA片段相关或有多强[12]。
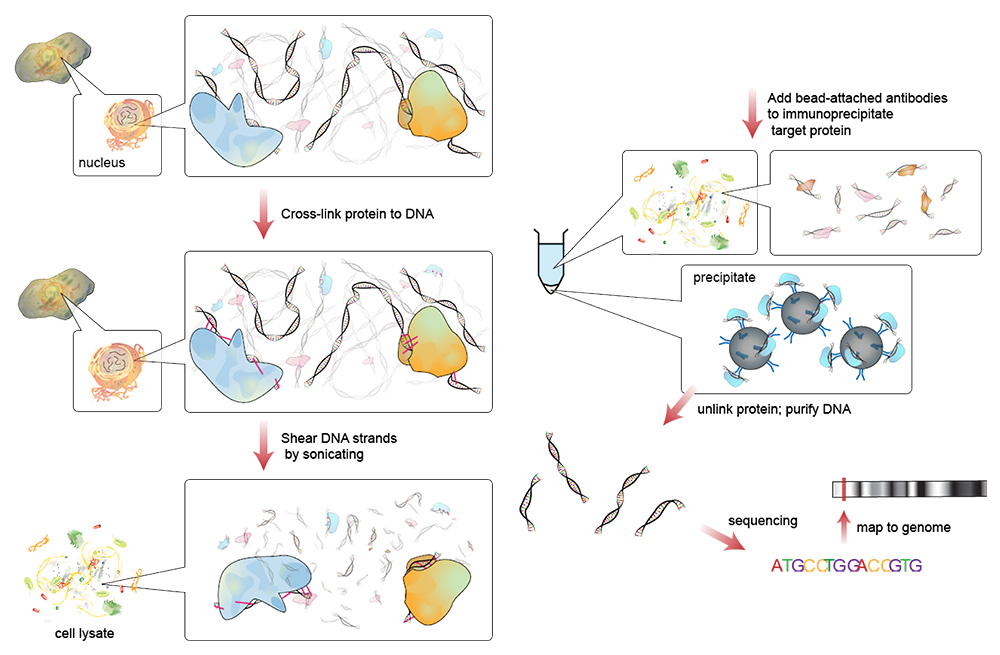
图2: 染色质免疫沉淀 (ChIP) 工作流程。 首先,DNA和相关蛋白质之间的结合是固定的。然后,通过超声将DNA切割成50至1000 bp长的片段。使用针对DNA相关靶蛋白的特异性抗体,选择性地免疫沉淀DNA-蛋白复合物。分离出的复合物后,可以纯化DNA并确定其序列[13]。
ChIP 中使用的抗体必须具有高质量才能使实验成功。我们的供应商 化验精灵 提供多种经过验证的优质 ChIP 抗体,适用于普通和新型 PTM。这些包括的修饰,如甲基化或乙酰化,以及相当奇特的修饰,如内地酰化或SUMO化[14]。
最著名和最重要的PTM之一是乙酰化。在这个过程中,乙酰基通过将其连接到氨基酸残基的氮原子来添加到蛋白质中。这种变化会对受影响的蛋白质产生深远的影响,例如其功能、稳定性或定位的改变[14]。最常见的乙酰化蛋白是组蛋白,其修饰仅发生在赖氨酸上。乙酰基的附着导致核小体构象的开放,使相应的基因可用于RNA聚合酶的转录[3]。因此,组蛋白乙酰化开启了某些基因的表达。
甲基化代表乙酰化的对应物,是最著名的表观遗传信号。除了组蛋白,DNA也可以直接甲基化。在此过程中,通过DNA甲基转移酶将甲基添加到胞嘧啶-鸟嘌呤序列(CpG)内的胞嘧啶核苷酸中[14]。新鲜甲基化的CpG导致所谓的阻遏蛋白的募集,其抑制DNA和转录因子之间的相互作用。结果,相应基因组片段中的基因表达受到抑制。在甲基化组蛋白的情况下,组蛋白构象是封闭的,因此基因的转录不再可能[3]。组蛋白甲基化存在于赖氨酸和精氨酸上[7]。
磷酸化描述了ATP的磷酸基团通过蛋白激酶添加到分子中的过程。反向反应称为去磷酸化,由蛋白质磷酸酶进行。组蛋白磷酸化可发生在具有羟基(丝氨酸、苏氨酸和酪氨酸)的氨基酸上。磷酸基团的插入显着增加了组蛋白的负电荷,从而改变了染色质的结构。组蛋白磷酸化功能多种多样,有助于基因表达的活化和失活[7]。
泛素化描述了泛素(一种由64个氨基酸组成的蛋白质)与另一种蛋白质的附着。这标志着蛋白酶体或溶酶体降解的目标蛋白。已知三种主要类型的泛素化:单泛素化和多泛素化以及多位点单泛素化。在单泛素化中,只有一个泛素分子连接到蛋白质上,而在多泛素化中,多个分子连接到单个蛋白质上。在多位点泛素化中,不止一个泛素分子连接到靶蛋白的单个赖氨酸残基上[14]。组蛋白的泛素化与真核基因表达的激活有关,但这种调控的分子基础在很大程度上仍然未知[15]。
在了解了四个最重要和最著名的 PTM 之后,我们现在转向更“异国情调"的修改。这些并不受欢迎,但仍然对所涉及的蛋白质有有趣的影响。这种异国情调的一个例子是β-羟基-丁酰化。这是酶将酮体β-羟基丁酸酯附着到组蛋白上的过程。这种修饰会影响结构,从而影响受影响的组蛋白的功能。β-羟基丁酰化与多种组蛋白相关疾病有关,因此可能是新疗法令人兴奋的靶点[14]。
SUMO化是一种PTM,其中所谓的SUMO蛋白与其他蛋白质的赖氨酸残基共价结合。相扑蛋白(s商场 u比素相关 莫difier)与泛素具有结构相似性,并在所有真核生物中形成高度保守的蛋白质家族。SUMO化在许多细胞过程中起重要作用,包括蛋白质-蛋白质相互作用、核细胞质转运、信号转导和细胞周期调节[16]。这种修饰也会影响蛋白质稳定性:SUMO化蛋白质通常比未修饰的蛋白质更稳定[14]。
Neddylation描述了一种PTM,其中泛素样蛋白NEDD8(神经-前体-细胞表达发育下调 8)与靶蛋白偶联。在此过程中,NEDD8的C端甘氨酸的羧基与靶蛋白中赖氨酸的Ɛ-氨基之间形成同肽键。该过程类似于泛素化,但涉及其他酶。Neddylization在各种细胞过程中起着至关重要的作用,例如转录,细胞接触和泛素 - 蛋白酶体系统的调节。内迪尔化失调可能导致各种疾病的发展,包括癌症、神经退行性疾病和心脏病[17]。
作为最后的PTM,让我们简要介绍一下巴豆酰化。在最近发现的修饰中,巴豆酰辅酶A分子连接到目标蛋白的赖氨酸残基上。这通常是组蛋白中赖氨酸的Ɛ-氨基基团。巴豆酰辅酶A是巴豆酸和辅酶A之间的硫酯。它在氨基酸L-赖氨酸和L-色氨酸的降解中作为代谢物被发现。巴豆酰化影响许多不同的蛋白质,包括组蛋白、转录因子和酶。它影响蛋白质的功能和稳定性。根据巴豆酰辅酶A分子的定位,靶蛋白的活性可以增加或减少。这种修饰也被怀疑对癌症等疾病的发展有影响[18]。
在这里,您将发现来自检测精灵的所有 ChIP 验证抗体: 来自检测精灵的 CHIP 验证抗体
表观遗传变化在一生中发生的频率是基因突变的很多倍[19]。因此,许多科学家确信,在未来,表观遗传学将为一些以前无法用遗传学解释的与年龄相关的疾病提供答案。因此,表观遗传学研究可以为创新疗法提供动力,从而为医学研究提供新的方向。
其基础是对表观遗传修饰的分析,例如组蛋白修饰。此处描述的翻译后修饰仅代表已知PTM的一小部分。 您想了解有关其他修改的更多信息吗?然后看看 本页 来自我们的供应商 化验精灵! 这里 您还将找到 Assay Genie 全面选择高度验证的染色质免疫沉淀抗体的概述。
我们的供应商 罗克兰免疫化学品 还为您的表观遗传学研究提供高质量的工具!为了检测不同的组蛋白修饰,这家美国公司通过多种特异性抗体支持您的研究,其中许多可用于染色质免疫沉淀:
| Product | Clonality | Reactivity | Application |
| Anti-acetyl-Lysine | polyclonal | broad | WB, IP |
| Anti-methylated Lysine | polyclonal | broad | WB |
| Anti-SUMO | polyclonal | broad | WB, ChIP, IP, ELISA |
| Anti-Ubiquitin | polyclonal | broad | WB, ELISA |
| Product | Clonality | Reactivity | Application |
| Anti-methyl-Histone H3 (R2me1) | polyclonal | human, C. elegans | WB, Dot Blot |
| Anti-dimethyl-Histone H3 (R2me2a) | polyclonal | human, C. elegans | WB, IF, Dot Blot |
| Anti-dimethyl-Histone H3 (R2me2s/K4me2) | polyclonal | human, mouse, C. elegans | WB, IF, ChIP, Dot Blot |
| Anti-acetyl-Histone H3 (K4ac) | polyclonal | human, mouse, C. elegans | WB, IF, ChIP, Dot Blot |
| Anti-monomethyl-Histone H3 (K4me1) | polyclonal | human | WB, Dot Blot, ELISA |
| Anti-dimethyl-Histone H3 (K4me2) | polyclonal | human, C. elegans | WB, IF, ChIP, Dot Blot |
| Anti-trimethyl-Histone H3 (K4me3) | polyclonal | human, mouse, C. elegans | WB, IF, ChIP, Dot Blot |
| Anti-phospho-Histone H3 (T6) | polyclonal | human, C. elegans | WB, IF, Dot Blot |
| Anti-dimethyl-Histone H3 (R8me2a) | polyclonal | human, mouse, C. elegans | WB, IHC, IF, ChIP, Dot Blot |
| Anti-dimethyl-Histone H3 (R8me2s) | polyclonal | human, C. elegans | WB, ChIP, Dot Blot |
| Anti-acetyl-Histone H3 (K9ac) | polyclonal | human, mouse, C. elegans | WB, IF, ChIP, Dot Blot |
| Anti-acetyl-Histone H3 (K9ac/K14ac) | polyclonal | human, mouse, C. elegans | WB, IF, Dot Blot |
| Anti-methyl-Histone H3 (K9me1) | polyclonal | human, mouse, C. elegans | WB, IF, ChIP, Dot Blot |
| Anti-dimethyl-Histone H3 (K9me2) | polyclonal | human, C. elegans | WB, IF, ChIP, Dot Blot |
| Anti-trimethyl-Histone H3 (K9me3) | polyclonal | human, C. elegans | WB, IF, ChIP, Dot Blot |
| Anti-phospho-Histone H3 (S10) | polyclonal | human, C. elegans | WB, IF, Dot Blot |
| Anti-phospho-Histone H3 (S10/T11) | polyclonal | human, C. elegans | WB, IF |
| Anti-phospho-Histone H3 (T11) | polyclonal | human | WB, IF, Dot Blot |
| Anti-acetyl-Histone H3 (K18ac) | polyclonal | human, mouse, C. elegans | WB, IF, ChIP, Dot Blot |
| Anti-methyl-Histone H3 (K18me1) | polyclonal | human, mouse | WB, IF, Dot Blot |
| Anti-dimethyl-Histone H3 (K18me2) | polyclonal | human, mouse, C. elegans | WB, IF, ChIP, Dot Blot |
| Anti-trimethyl-Histone H3 (K18me3) | polyclonal | human, mouse | WB, IF |
| Anti-acetyl-Histone H3 (K23ac) | polyclonal | human, mouse | WB, Dot Blot |
| Anti-dimethyl-Histone H3 (K23me2) | polyclonal | human, mouse, rat, C. elegans | WB, ELISA |
| Anti-acetyl-Histone H3 (K27ac) | polyclonal | human, mouse, C. elegans | WB, ChIP, Dot Blot |
| Anti-methyl-Histone H3 (K27me1) | polyclonal | human | WB, IF, ChIP, Dot Blot, ELISA |
| Anti-dimethyl-Histone H3 (K27me2) | polyclonal | human | WB, IF, ChIP, Dot Blot, ELISA |
| Anti-trimethyl-Histone H3 (K27me3) | polyclonal | human, mouse, rat | WB, IF, Dot Blot |
| Anti-phospho-Histone H3 (S28) | polyclonal | human, mouse | WB, IF |
| Anti-acetyl-Histone H3 (K36ac) | polyclonal | human, C. elegans | WB, Dot Blot |
| Anti-methyl-Histone H3 (K36me1) | polyclonal | human, C. elegans | WB, IF, Dot Blot |
| Anti-dimethyl-Histone H3 (K36me2) | polyclonal | human, mouse, C. elegans | WB, IF, ChIP, Dot Blot |
| Anti-trimethyl-Histone H3 (K36me3) | polyclonal | human, mouse, C. elegans | WB, IF, Dot Blot |
| Anti-methyl-Histone H3 (K37me1) | polyclonal | human, mouse, C. elegans | WB, IF, Dot Blot |
| Anti-dimethyl-Histone H3 (K37me2) | polyclonal | human, mouse, C. elegans | WB, IF, Dot Blot |
| Anti-trimethyl-Histone H3 (K37me3) | polyclonal | human, C. elegans | WB, IF, Dot Blot |
| Anti-methyl-Histone H3 (K56me1) | polyclonal | human, mouse | WB, IF, Dot Blot |
| Anti-trimethyl-Histone H3 (K56me3) | polyclonal | human, C. elegans | WB, IF, Dot Blot |
| Anti-methyl-Histone H3 (K79me1) | polyclonal | human, mouse, monkey | WB, IF, ChIP, Dot Blot |
| Anti-trimethyl-Histone H3 (K79me3) | polyclonal | human, C. elegans | WB, IF, ChIP, Dot Blot |
| Product | Clonality | Reactivity | Application |
| Anti-phospho-Histone H4 (S1) | polyclonal | human | WB, IF, ChIP, Dot Blot |
| Anti-methyl-Histone H4 (R3me1) | polyclonal | human, mouse | WB, IF |
| Anti-acetyl-Histone H4 (K5ac) | polyclonal | human, mouse | WB, IF, ChIP |
| Anti-acetyl-Histone H4 (K8ac) | polyclonal | human, mouse, C. elegans | WB, IF, ChIP, Dot Blot |
| Anti-acetyl-Histone H4 (K12ac) | polyclonal | human | WB, IF, Dot Blot, Microarray |
| Anti-acetyl-Histone H4 (K16ac) | polyclonal | human, mouse | WB, IF, Dot Blot |
| Anti-methyl-Histone H4 (K20me1) | polyclonal | human, mouse, C. elegans | WB, IF, ChIP, Dot Blot |
| Anti-dimethyl-Histone H4 (K20me2) | polyclonal | human, C. elegans | WB, IF, ChIP, Dot Blot |
| Product | Clonality | Reactivity | Application |
| Anti-Histone H2 A.Zac | polyclonal | broad | WB, IF, ChIP, Dot Blot, ELISA |
| Anti-phospho-Histone H2AvD (S137) | polyclonal | D. melanogaster | WB, IHC, IF, EM, ELISA |
| Anti-phospho-H2AX (S139) | polyclonal | human | WB, ELISA |
 扫一扫,关注微信
扫一扫,关注微信 当前位置:
当前位置: